2. F-class ion pumps:
The F class ion pumps contain different transmembrane and cytosolic subunits. They are known for only transport of protons, in a process that does not involve phosphoprotein intermediate. They generally behave as reverse proton pump by synthesizing ATP from ADP and Pi by movement of protons from the exoplasmic to the cytosolic face of the membrane down the proton electrochemical gradient. Therefore, these pumps are also known as ATP synthases or F0F1 complex. F-class ion pump is most common in bacteria, yeast and animal mitochondria and also in chloroplast.
The F0F1 complex is a multi-protein having two components F0and F1. Both are multimeric proteins. The F0 component contains three integral membrane proteins named a, b and c. The a and two b subunits are linked tightly but not to the donut- shaped ring of c subunits. And the F1 component is water soluble complex of five distinct polypeptides with the composition α3β3γδε. The lower part of the F1γ subunit is a coil which fits into the centre of the c-subunit ring of F0 and appears rigidly attached to it. The F1 ε subunit is rigidly attached to γ and also forms rigid contacts with c subunits. The F1 and subunits associate in alternating order to form a hexamer αβαβαβ. The F1δ subunit is permanently linked to one of the F1 subunits and also to the b subunit of F0. Thus the F0 a and b subunits and the δ subunit and (αβ)3 hexamer of the F1 complex form a rigid structure anchored in the membrane. The rodlike b subunits form a stator that prevents the (αβ)3 hexamer from moving while it rests on the γ subunit.
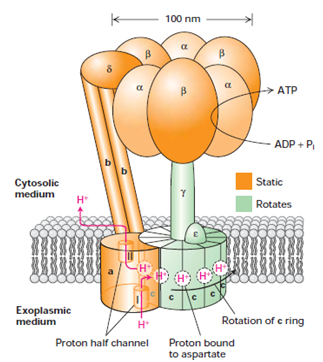
Figure 2: Model of the structure and function of ATP synthase (the F0F1 complex) in the bacterial plasma membrane. The F0 portion is built of three integral membrane proteins: one copy of a, two copies of b, and on average 10 copies of c arranged in a ring in the plane of the membrane. Two proton half-channels lie at the interface between the a subunit and the c ring. Half-channel I allows protons to move one at a time from the exoplasmic medium and bind to aspartate-61 in the center of a c subunit near the middle of the membrane. Half-channel II (after rotation of the c ring) permits protons to dissociate from the aspartate and move into the cytosolic medium. [Adapted from M. J. Schnitzer, 2001, Nature 410:878, and P. D. Boyer, 1999, Nature 402:247.]