Sodium:
Na+ accumulation is accompanied by vacuolar alkalinization in barley roots. This was established by using NMR spectroscopy.
Calcium:
Calcium plays a central role in signal transduction and higher concentrations are observed in apoplast and within the vacuole. An energized, highly specific calcium uptake by the vacuole is, therefore, a prerequisite for maintaining a low cytosolic calcium concentration. P-type Ca-ATPases have been identified at the plasma membrane, the ER, and the vacuolar membrane. A Ca2+ pump called a Ca2+/H+ antiporter has been demonstrated in vacuolar membrane fractions. This antiporter exhibits a far lower affinity than the Ca2+-ATPase. Also, a vacuolar voltage gated Ca2+ channel (VVCa) has been reported which is activated on membrane hyperpolarization.
Magnesium:
The presence of a Mg2+/H+ antiporter has been described for the vacuole-like lutoids of Hevea brasiliensis and tonoplast vesicles isolated from maize roots.
Heavy metals:
Plants need some heavy metals such as Cu2+ or Zn2+ as micronutrients. Therefore, they need to be transported and a large portion of the heavy metals absorbed by the cell is usually concentrated within the vacuole. A vacuolar Cd2+/H+ antiport activity has been demonstrated. However, it is known that plants form chelates with heavy metals by synthesizing phytochelatins (PCs), and these PCs can be transported into vacuoles of Schizosaccharomyces as apoPC or as PC-Cd complexes by ABC transporters. Vacuoles of higher plants are also known to transport phytochelatins.
Transport of products of the secondary metabolites:
Involvement of secondary energized transporters and directly energized, ABC-type transporters
Plants synthesize an huge number of secondary metabolites and many of these have been found to be exclusively localized in the vacuole. The electrochemical gradient established by the two vacuolar proton pumps is used by the secondary energized transporters as a source of energy. It was demonstrated that the ΔpH was essential for the uptake of a number of phenolics, such as esculin, o-coumaric acid glucoside, apigenin- 7-(6-O-malonyl) glucoside, and anthocyanins from carrot. Recently it became evident that in addition to transporters depending on the proton motive force, directly energized transporters are also present on the vacuolar membrane. The first demonstration for a directly activated transport of solutes into the vacuole was provided for glutathione conjugates. Flavonoid glucuronides, a secondary plant compounds in rye vacuoles are transported by directly energized transport processes. Furthermore, studies with lucifer yellow, a sulfonated compound also indicates that sulfonated and sulfated secondary compounds cross the tonoplast by direct energization.
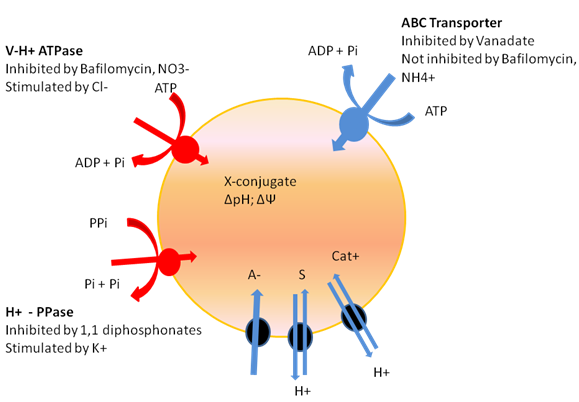
Figure 7: Proton pumps establishing a electrochemical gradient (red), secondary energized uptake mechanisms (green), and directly energized, ABC-type transporters (blue) of the plant vacuole. S, neutral solute; A–, anion; cat+, cation; X-conjugate, conjugate of a compound X (secondary metabolite or xenobiotic) with a hydrophilic compound such as glucose, glutathione, an amino acid, malonate, or sulphate.
Interesting facts:
- During endocytosis, these intra-lysosomal membranes are formed and prepared for digestion by a lipid-sorting process during which their cholesterol content decreases and the concentration of the negatively charged bis(monoacylglycero)phosphate increases.
- Lysosomal enzyme disorders contribute to several human diseases, either due to genetic defects in its enzyme expression or the escape of lysosomal enzymes (lysozymes) into extralysosomal medium.
- Permeabilization of lysosome, has been shown to initiate a cell death pathway or apoptosis.