Euchromatin: The lightly-stained regions in chromosome when stained with basic dyes are called euchromatin and contain single-copy of genetically-active DNA. The extent of chromatin condensation varies during the life cycle of the cell and plays an important role in regulating gene expression. In the interphase of cell cycle the chromatin are decondensed and known as euchromatin leading to gene transcription and DNA replication.
Heterochromatin: The word heterochromatin was coined by Emil Heitz based on cytological observations. They are highly condensed and ordered areas in nucleosomal arrays. About 10% of interphase chromatin is called heterochromatin and is in a very highly condensed state that resembles the chromatin of cells undergoing mitosis. They contain a high density of repetitive DNA found at centromeres and telomeres form heterochromatin. Heterochromatin are of two types, the constitutive and facultative heterochromatin. The regions that remain condensed throughout the cell cycle are called constitutive heterochromatin whereas the regions where heterochromatin condensation state can change are known as facultative. Constitutive heterochromatin is found in the region that flanks the telomeres and centromere of each chromosome and in the distal arm of the Y chromosome in mammals. Constitutive heterochromatin possesses very few genes and they also lead to transcriptional inactivation of nearby genes. This phenomenon of gene silencing is known as “position effect”. Constitutive heterochromatin also inhibits genetic recombination between homologous repetitive sequences circumventing DNA duplications and deletion. Whereas facultative heterochromatin is chromatin that has been specifically inactivated during certain phases of an organism’s life or in certain types of differentiated cells. Dosage compensation of X-chromosome or X-chromosome inactivation in mammals is an example of such heterochromatin (Karp 2010). Heterochromatin spreads from a specific nucleation site, causing silencing of most of the X chromosome, thereby regulating gene dosage.
Centromeres: Centromeres are those condensed regions within the chromosome that are responsible for the accurate segregation of the replicated chromosome during mitosis and meiosis. When chromosomes are stained they typically show a dark-stained region that is the centromere. The actual location where the attachments of spindle fibres occur is called the kinetochore and is composed of both DNA and protein. The DNA sequence within these regions is called CEN DNA. Because CEN DNA can be moved from one chromosome to another and still provide the chromosome with the ability to segregate, these sequences must not provide any other function. Typically CEN DNA is about 120 base pairs long and consists of several sub-domains, CDE-I, CDE-II and CDE-III (Figure 3). Mutations in the first two sub-domains have no effect upon segregation, but a point mutation in the CDE-III sub-domain completely eliminates the ability of the centromere to function during chromosome segregation. Therefore CDE-III must be actively involved in the binding of the spindle fibers to the centromere. The protein component of the kinetochore is only now being characterized. A complex of three proteins called Cbf-III binds to normal CDE-III regions but cannot bind to a CDE-III region with a point mutation that prevents mitotic segregation. Furthermore, mutants of the genes encoding the Cbf-III proteins also eliminates the ability for chromosomes to segregate during mitosis. Additional analyses of the DNA and protein components of the centromere are necessary to fully understand the mechanics of chromosome segregation.
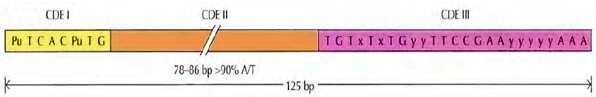
Figure 3: The S. cerevisiae centrosome. The S. cerevisae centromere (CEN) sequences consist of two short conserved sequences (CDE I and CDE Ill) separated by 78 to 86 base pairs (bp) of ATrich DNA (CDE II). The sequences shown are consensus sequences derived from analysis of the centromere sequences of individual yeast chromosomes. Pu = A or G; x =A or T; y = any base. The figure has been adapted from “The Cell, A Molecular Approach” by Geoffrey M. Cooper, 4th Ed. 2007.