Structure of nuclear pores: Nuclear pores appear circular in surface view and have a diameter between 10nm to 100 nm. Previously it was believed that a diaphragm made of amorphous to fibrillar material extends across each pore limiting free transfer of material. Such a diaphragm called annulus has been observed in animal cells, but lack in plant cells. Recent electron microscopic studies have revealed that a nuclear pore has far more complex structure, so it is called nuclear pore complex with an estimated molecular weight of 50 to 100 million daltons. Negative staining techniques have demonstrated that pore complexes have an eight-fold or octagonal symmetry.
Nuclear Pore density: In nuclei of mammals it has been calculated that nuclear pores account for 5 to 15 per cent of the surface area of the nuclear membrane. In amphibian oocytes, certain plant cells and protozoa, the surface occupied by the nuclear pores may be as high as 20 to 36 per cent.
Arrangement of nuclear pores on nuclear envelope: In somatic cells, the nuclear pores are
evenly or randomly distributed over the surface of nuclear envelope. However, pore arrangement in other cell types is not random but rather range from rows (spores of Eqisetum) to Clusters (oocytes of Xenopus laevis) to hexagonal (Malpighian tubules of leaf hoppers) packing order.
Nucleo-cytoplasmic traffic: Quite evidently there is considerable trafficking across the nuclear envelope during interphase. Ions, nucleotides and structural, catalytic and regulatory proteins are imported from the cytosol (cytoplasmic matrix); mRNA, tRNA are exported to the cytosol (cytoplasmic matrix). However, one of the main functions of the nuclear envelope is to prevent the entrance of active ribosomes into the nucleus.
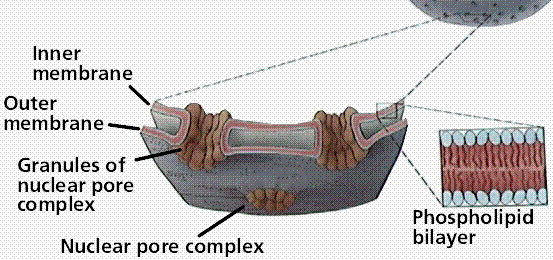
Figure 2: An illustration of the nuclear membrane
Nucleoplasm:
The space between the nuclear envelope and the nucleolus is filled by a transparent, semi-solid,
granular and slightly acidophilic ground substance or the matrix known as the nuclear sap or
nucleoplasm or karyolymph. The nuclear components such as the chromatin threads and the
nucleolus remain suspended in the nucleoplasm which is composed mainly of nucleoproteins
but it also contains other inorganic and organic substances, namely nucleic acids, proteins, enzymes and minerals. The most common nucleic acids of the nucleoplasm are the DNA and RNA. The nucleoplasm contains many types of complex proteins
categorized into: (i) Basic proteins. The proteins which take basic stain are known as the basic proteins. The most important basic proteins of the nucleus are nucleoprotamines and the nucleohistones. (ii) Non-histone or Acidic proteins. The acidic proteins either occur in the nucleoplasm or in the chromatin. The most abundant acidic proteins of the euchromatin (a type of chromatin) are the phosphoproteins. The nucleoplasm contains many enzymes which are
necessary for the synthesis of the DNA and RNA. Most of the nuclear enzymes are composed of non-histone (acidic) proteins. The most important nuclear enzymes are the DNA polymerase, RNA polymerase, NAD synthetase, nucleoside triphosphatase, adenosine diaminase, nucleoside phosphorylase, guanase, aldolase, enolase, 3-phosphoglyceraldehyde dehydrogenase and pyruvate kinase. The nucleoplasm also contains certain cofactors and coenzymes such as ATP and acetyl CoA. The nucleoplasm has small lipid content. The nucleoplasm also contains several inorganic compounds such as phosphorus, potassium, sodium, calcium and magnesium. The chromatin comparatively contains large amount of
these minerals than the nucleoplasm.
The nucleoplasm contains many thread-like, coiled and much elongated structures which take readily the basic stains such as the basic fuchsin. These thread-like structures are known as the chromatin (chrome=colour) substance or chromatin fibres. Chromosome will be discussed in detail in the next module.